Доктор
БАСОВ
#fascia #manualtherapy #osteopathy #yogatherapy #massage #фасция #остеопатия #йогатерпаия #массаж #докторбасов
Мануальная терапия кусочка свежего мяса: что здесь не так?
Труп это тело минус жизнь. Хотя с этим утверждением легко согласиться, связанный с ним более интригующий вопрос включает в себя:
• Как именно «жизнь» влияет на реакцию тканей во время мануальной терапии?
• Какие физиологические или другие известные механизмы участвуют в этом?
• Могут ли ответы на этот вопрос также помочь понять разницу между качествами тканей в, казалось бы, очень живых частях тела и в более запущенных частях тела, которые ощущаются менее живыми для пальпирующей руки?
Руководствуясь этими вопросами, автор и группа его коллег провели несколько исследовательских экспериментов, в которых они применяли методы ручной мобилизации для свежеиссеченной фасции животных (поясничная фасция свиней, коров, крыс и мышей, взятых через несколько минут после их умерщвления; см. Рис.1). Хотя было возможным ослабить некоторые из более мелких коллагеновых соединений и спаек, у практикующих осталось четкое впечатление, что многие важные аспекты реакции ткани, которые обычно ощутимы при работе с «живой фасцией», отсутствовали в этих сеансах искусственного лечения. В аналогичном эксперименте автор применил методы фасциальной мобилизации к пациентам, находящимся под наркозом в условиях больницы. Здесь также наблюдается отсутствие реакции нормальной ткани. В то время как многие ранее напряженные мышечные ткани (такие как верхняя трапеция или подколенные сухожилия) оказались более мягкими по сравнению с их состоянием до анестезии, ни одна из реакций плавления или релиза, знакомые фасциотерапевтам, не была обнаружена во время применения методов глубокой мобилизации тканей (Schleip, 2012).
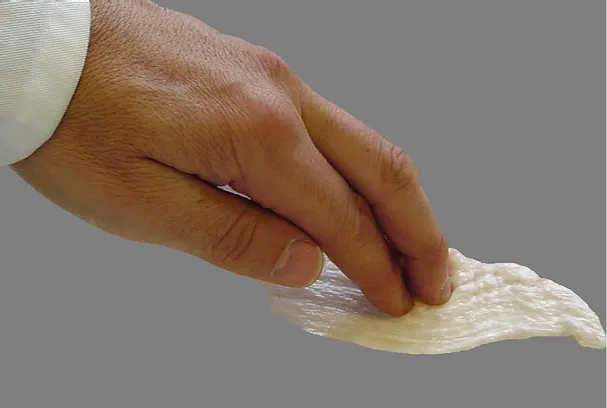
Рисунок 1. Эксперименты с различными миофасциальными техниками на свежем куске животной фасции. Хотя большинство врожденных фибробластных клеток все еще живы (в течение нескольких часов после умерщвления донора), некоторые тканевые реакции, знакомые фасциотерапевтам при работе с фасцией в нормальных условиях, не могут быть получены. Может ли это отсутствие ответа быть связано с нарушением нервных связей с центральной нервной системой?
Хотя эти исследования не включали в себя достаточную методологическую строгость и, следовательно, на них нельзя полагаться с научной точки зрения, они, похоже, согласуются с результатами обширного исследования, проведенного Chaudhry et al. (2008). Здесь было показано, что механические силы, используемые при миофасциальной мобилизации, способны вызывать немедленную и длительную деформацию ткани в очень рыхлых соединительных тканях, таких как носовая фасция. Однако этих сил недостаточно, чтобы вызвать длительные эффекты в плотных фасциальных тканях (таких как поясничная фасция, фасция lata или подошвенная фасция). Поэтому авторы исследования предположили, что фасциальная терапия с ощутимыми результатами может включать стимуляцию сенсорных нервных окончаний в фасции, которая через их связь с центральной нервной системой может вызывать физиологические или нервно-мышечные реакции, которые косвенно влияют на восприимчивость тканей (помимо первоначального эффекта биомеханической деформации ткани в этом исследовании).
Фасциальные ткани богато иннервируются.
Что на самом деле известно о сенсорной иннервации фасции? Фактически, для большинства медицинских работников фасция долгое время считалась инертным обертывающим органом, обеспечивающим механическую поддержку другим более важным структурам, таким как мышцы и органы. В то время как существовали некоторые ранние гистологические отчеты о наличии сенсорных нервов в фасции (Stillwell, 1957; Sakada, 1974), они были едва замечены и не влияли на общее понимание динамики скелетно-мышечной системы. В то время как другие пионеры мануальной терапии, такие как Моше Фельденкрайз, а также Ида Рольф, основатели соматической терапии, очевидно, не знали о важности фасции как сенсорного органа, именно основатель остеопатии, Эндрю Тейлор Стилл, провозгласил что «Без сомнения, нервы существуют в фасции ...» и предложил, чтобы ко всем фасциальным тканям относились с такой же степенью уважения, как если бы они имели дело с «ответвлениями мозга». (Still, 1902 г.) Однако его труды не содержали никаких подробностей о том, как он пришел к такому выводу, какие конкретно типы нервов он имел в виду, и как они реагируют на различные виды механической стимуляции. Большой вклад в эту область позже дал Ван дер Валь, который с кропотливой детальностью задокументировал богатое присутствие сенсорных нервных окончаний в фасции крыс (Van der Wal, 1988; Van der Wal, 2009). К сожалению, это важное открытие едва ли было замечено медицинским и терапевтическим сообществом в течение нескольких десятилетий. Относительно связок - как очень специализированных фасциальных тканей - их проприоцептивная иннервация была признана в течение 1990-х годов, что впоследствии повлияло на рекомендации по ортопедическим операциям (Johansson et al., 1991). Аналогично, было обнаружено, что подошвенная фасция как специализированная ткань способствует сенсомоторной регуляции постурального контроля в положении стоя (Erdemir & Piazza, 2004). Тем не менее, что действительно изменило общую перспективу в более всеобъемлющей и мощной манере, так это первый международный конгресс исследований фасции, проведенный в Гарвардской медицинской школе в Бостоне в 2007 году. Во время Конгресса три команды из разных стран независимо друг от друга сообщили о своей находке - обилие чувствительных нервов в фасциальных тканях (Findley & Schleip, 2007).
На рис. 2 приведен пример богатого представительства нервной системы в фасции человека. Если признавать все преимущественно волокнистые соединительные ткани - такие как связки, капсулы органов или внутримышечные соединительные ткани - как часть растягивающей сети всего тела, называемой «фасция», то фасция содержит по крайней мере, несколько сотен тысяч нервных окончаний, которые соединяют периферические фасциальные ткани с центральной соматической или автономной нервной системой (Mitchell & Schmidt, 1977).
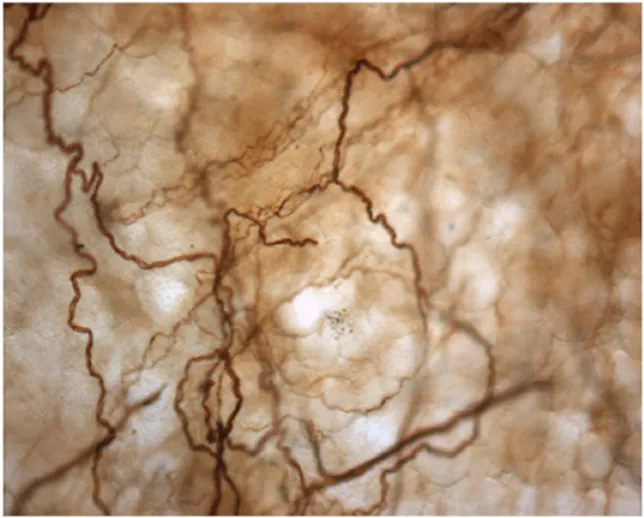
Рисунок 2. Демонстрация обильного присутствия нервов в кусочке поясничной фасции человека. Длина изображения составляет примерно 0,5 мм. Использование паннейрональных антител позволило сделать видимой маркировку всех нервных тканей (здесь темно-коричневого цвета). Воспроизводится по Tesarz J., Hoheisel U., Wiedenhöfer B., Mense S. (2011). Сенсорная иннервация грудопоясничной фасции у крыс и людей. Neuroscience 194: 302–308
В свою очередь, фасция снабжается богатым запасом эфферентных симпатических нервных окончаний, которые влияют на местную вазомоторную активность и могут иметь дополнительные (пока не понятные) функции (Tesarz et al, 2011). Исходя из этого, кажется не слишком самонадеянным смотреть на фасциальную сеть всего тела как на сенсорный орган. Что касается огромного количества и богатства нервных окончаний, то лицевая сеть может «противостоять» нашим ощущениям зрения, не говоря уже о слухе или любом другом из наших обычно рассматриваемых органов чувств. Когда запутанная система внутримышечных мешочков и перегородок коллагеновых соединительных тканей включена в качестве способствующих элементов этой натяжной сети, тогда фасция также может рассматриваться как наш самый большой орган чувств с точки зрения общей площади ее поверхности. Согласно Tesarz и соавт. (2011) характер иннервации фасции, исследованный на грызунах и поясничной фасции человека, следует сегментарному типу, сопоставимому с конгруэнцией хорошо известных карт иннервации миотомов и дерматомов. Термин «фасциатома» был предложен для фасциальных областей, иннервируемых соответствующими уровнями спинного мозга.
Участки с особенно богатой иннервацией.
Имеет ли значение, какие места фасциальной сети стимулировать при мануальной терапии, для того чтобы обеспечить спинной мозг новой проприоцептивной информацией? Два новых взгляда на плотность сенсорных рецепторов в фасции дают ценное понимание этого вопроса. Во-первых, недавние гистологические исследования группы Mense в Гейдельбергском университете показали, что как в поясничной фасции человека, так и в области поясницы у грызунов плотность сенсорных нейронов значительно выше в самых поверхностных слоях ткани, то есть в тех, что находятся между дермой и глубокой фасцией ( Tesarz et al., 2011). В наших собственных экспериментах в Ульмском университете мы также наблюдали увеличение плотности видимых нервов в переходной зоне сдвига между глубокой и поверхностной фасцией. В здоровых областях тела в этой зоне можно легко вызвать боковое «скольжение кожи» по отношению к нижележащим тканям. Это также зона, архитектура которой определяет, можно ли отделить кожную складку от тела или нет. Это может послужить основой для правдоподобного предположения, что боковые скользящие движения, обеспечиваемые повседневными движениями, являются важным источником проприоцепции фасции. В связи с этим, также можно предположить, что часто глубокие анекдотические эффекты различных методов тейпирования кожи в спортивной медицине могут быть частично объяснены локальным усилением соответствующих движений кожи при нормальном функционировании суставов. Для подтверждения этих предположений необходимы дальнейшие исследования. Второе недавнее понимание пришло от группы Stecco в Университете Падуи в Италии (Stecco et al., 2007). Их гистологические исследования фасций верхних и нижних конечностей у трупов человека выявили огромные различия в плотности проприоцептивных нервных окончаний, таких как тельца Гольджи, Пачини и Руффини. Их данные указывают на то, что фасциальные ткани, которые явно выполняют важную функцию передачи силы (например, lacertus fibrosus на верхнем предплечье как продолжение двуглавой мышцы плеча), почти не содержат проприоцептивных окончаний. С другой стороны, исследователи заметили, что другие фасциальные структуры, по-видимому, играют очень незначительную роль в передаче силы, о чем свидетельствует их отрезание, как в случае с фасцией вокруг лодыжки и запястья. Интересно, что эти более наклонно бегущие фасциальные тяжи, кажется, расположены на специфически коротких расстояниях от главных суставов, и содержат очень высокую плотность проприоцептивных нервных окончаний. Было даже высказано предположение, что основной функцией этих фасциальных групп может быть не их биомеханическая, а их сенсорная функция в обеспечении детальной проприоцепции центральной нервной системы. Если это подтвердится, это может указывать на то, что подходы, усиливающие проприоцепцию в мануальной терапии, могут быть увеличены в их соответствующей терапевтической эффективности путем стимуляции фасциальных тканей в областях с повышенной проприоцептивной иннервацией.
Различные типы механосенсорных рецепторов и их функции.
Рисунок 3 иллюстрирует типичный состав скелетно-мышечного нервного пучка, такого как седалищный или лучевой нерв. Одним из интересных аспектов этой композиции является то, что огромное часть нервных волокон выполняет функцию тонкого регулирования доставки питательных веществ через сосудистую систему, которая управляется симпатической нервной системой.
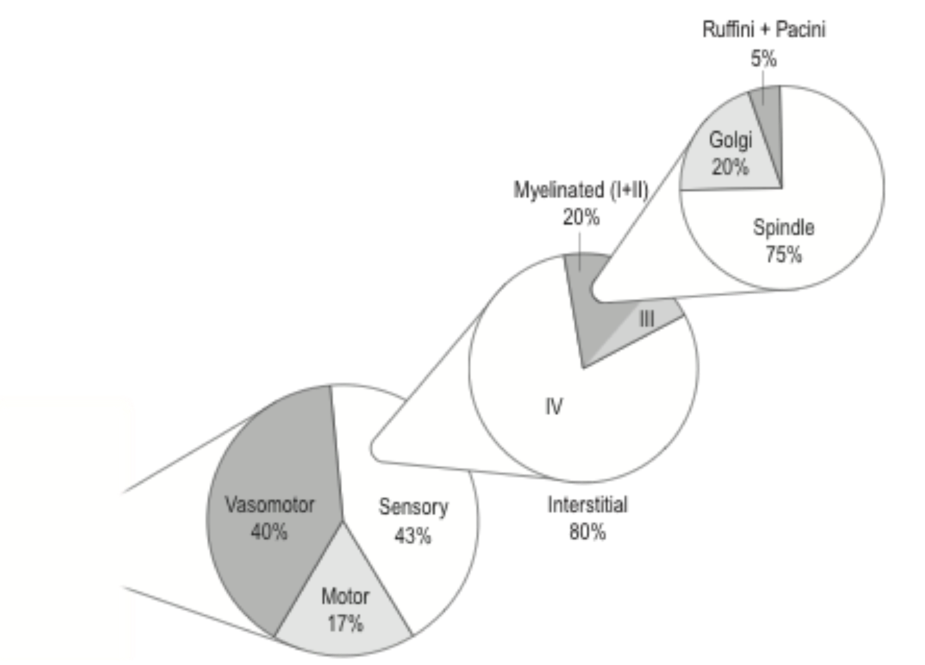
Оставшаяся часть, относящаяся к сенсомоторной регуляции, в совсем не одинаково разделена на моторные и сенсорные пути. Напротив, архитектура тела выделяет более чем в два раза больше нейронов для «слушания» или восприятия, чем для «разговора» и направления указаний к периферии. Может ли этот мудрый архитектурный принцип стать одним из объяснений того, почему иногда врожденный интеллект тела намного превосходит эффективность старомодного властного руководителя, который пытается руководить своей компанией с помощью множества требований и приказов с едва развитыми навыками слушания? Четверть сенсорных аксонов состоят из относительно быстро проводящих миелиновых нейронов, которые явно можно рассматривать как проприоцептивные пути. Их окончание включает тельца Пачини, Гольджи и Руффини. Они обычно оканчиваются в фасциальных тканях, либо в эпимизальной или сухожильной частях, либо во внутримышечных соединительных тканях.
Обратите внимание, что мышечные веретена - которые с эволюционной точки зрения являются довольно недавним изобретением для тонкого управления движения у наземных животных - могут рассматриваться как сенсорные приспособления, которые также расположены в перимизиальных или эндомизиальных коллагеновых тканях внутримышечной фасциальной сети. Вероятно, что если окружающие коллагеновые ткани страдают от потери эластичности, то функционирование этих веретен может быть нарушено. Возможно, это может быть фактором, способствующим возникновению таких состояний, как хроническая ригидность мышц или фибромиалгия, так как было показано, что увеличение толщины эндомизия характерно для пациентов с фибромиалгией (Liptan, 2010). Кроме того, от ученых изучающих мышечную ткань (мясо) также интересно узнать, что увеличение толщины перимизиума объясняет разницу в жесткости между жесткими и мягкими мышцами (жестким и мягким мясом) у одного и того же животного (Schleip et al., 2006). В то время как много известно о функционировании этих различных механочувствительных нейронов, большинство сенсорных нервов в фасциальной ткани заканчиваются интерстициальными свободными нервными окончаниями (см. Рис. 3), которые гораздо менее понятны и до сих пор считаются интригующе загадочными для тех, кто их изучает. Классическая неврология далее делит их на нейроны типа III, аксоны которых содержат очень тонкий слой миелина, и нейроны типа IV с немиелинизированными аксонами. Для наших целей, однако, можно считать, что они ведут себя аналогичным образом, хотя скорость проведения нейронов типа IV (также называемых С-волокнами в другой классификации) даже ниже, чем у и так медленных аксонов типа III (также называемых А-дельта волокна). Для обычных миелинизированных нейронов - снабжающих мышечные веретена, окончания Гольджи, Пачини и Руффини - ясно, что они функционируют как проприоцептивные устройства. Сообщалось, что эти окончания Руффини очень чувствительны к сдвиговым нагрузкам (то есть разница в направлении растягивающей нагрузки между одним слоем ткани и соседним); в то время как окончания Pacini реагируют только на быстрые изменения, и они имеют свойство быть невосприимчивыми к любым постоянным стимулам.
Хотя ранее считалось, что рецепторы Гольджи существуют только в сухожильных тканях, их присутствие в других фасциальных тканях было независимо подтверждено различными международными исследованиями (Yahia et al., 1992; Stecco et al., 2007; Stecco et al., 2008) , Стимуляция рецепторов Гольджи имеет свойство вызывать релаксационный ответ в тех мышечных волокнах, которые последовательно связаны с пучками стимулированных коллагеновых волокон. Однако если сухожильные ткани растягиваются с последовательно соединенными миофибриллами в расслабленном состоянии, то большая часть соответствующего удлинения будет «проглатываться» более податливыми миофибрилами. Представьте себе веревку, состоящую из плотной нити соединенной в линию с мягкой эластичной нитью. При вытягивании обоих, сначала происходит удлинение в более мягкой части, в то время как в последовательно соединенной более плотной части вряд ли будет видно какое-либо удлинение. (Вот почему хатха-йога может быть настолько эффективна в достижении расслабления. Пребывание в статической асане включает и приводит в тонус соответствующие мышцы, позволяя перенести растягивающую нагрузку на их сухожилия). Это соответствует статье Джами, в которой говорится, что растягивающая нагрузка с одновременно расслабленными мышцами может не обеспечивать достаточную стимуляцию для того, чтобы вызвать какое-либо изменение мышечного тонуса (Jami, 1992). Сообщалось, что стимуляция окончаний Руффини вызывает снижение симпатического влияния. Окончания Руффини кажутся особенно чувствительными к нагрузке сдвига. Это интригующая мысль, что медленные, плавящие миофасциальные методы, включающие навыки «локального слушания» относительно предпочтительного направления высвобождения ткани, могут быть особенно эффективными в стимуляции этих рецепторов. Такие методы часто используются в методе Рольфинга (Jones, 2004), а также в подходах, связанных с глубокими манипуляциями с тканями. Тельца Пачини с другой стороны предпочитают быстрые изменения, поскольку они являются быстроадаптируемыми рецепторами. Они могут быть особенно вовлечены в манипуляции с высокой скоростью, но также и в более мягких качающих движениях, которые используются в подходе Трагера или в Гармонической технике Ледермана (Trager, 1987; Schleip, 2003; Lederman, 2005). Было показано, что стимуляция механосенсорных интерстициальных свободных нервных окончаний, особенно с высоким механическим порогом, индуцирует изменение локального обмена жидкости посредством увеличения локальной вазодилатации, а также экструзии плазмы (Mitchell & Schmidt, 1977).
Рисунок 3. Нейронный состав периферического нерва. Проиллюстрированные пропорции были получены из количественного анализа отдельных аксонов, которые составляют больший нервный пучок, снабжающий периферическую область (в этом случае объединенный нерв, снабжающий боковые икроножные и подошвенные мышцы кошки; данные взяты из Mitchell & Schmidt, 1977) , В то время как небольшая часть интерстициальных нейронов может оканчиваться внутри кости, все остальные нейроны могут считаться оканчивающимися в фасциальных тканях. Даже сенсорные комплексы, называемые мышечными веретенами, расположены в перимизиальных или эндомизиальных фасциальных тканях. Интерстициальные нейроны оканчиваются свободными нервными окончаниями. Некоторые из них явно имеют проприоцептивную, интероцептивную или ноцицептивную функцию. Недавние исследования, однако, предполагают, что большинство интерстициальных нейронов в фасции выполняют полимодальную функцию, что означает, что они открыты для ноцицептивной, а также для неноцицептивной стимуляции.
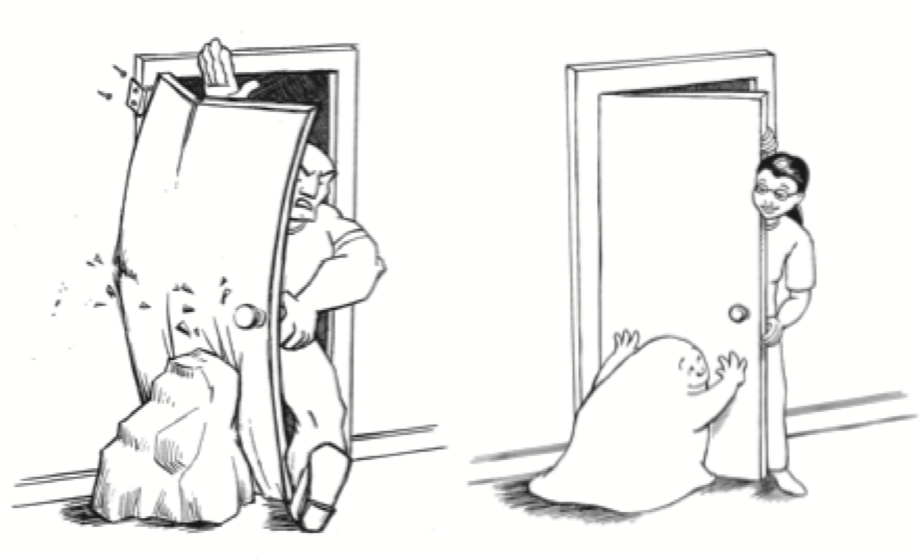
Рисунок 4. Два способа открыть заблокированную дверь или сустав. Дверь, заблокированная механическим объектом, требует другого подхода по сравнению с дверью, заблокированной живым препятствием. Точно так же к заблокированному суставу или неподвижной ткани можно приблизиться с чисто механической точки зрения, как будто речь идет о мертвых тканях. В качестве альтернативы к нему можно подойти как к механизму с активной саморегуляцией. Выбор подхода во многом зависит от того, осознает ли практикующий врач какую-либо нервную динамику, связанную с конкретной ситуацией у пациента.
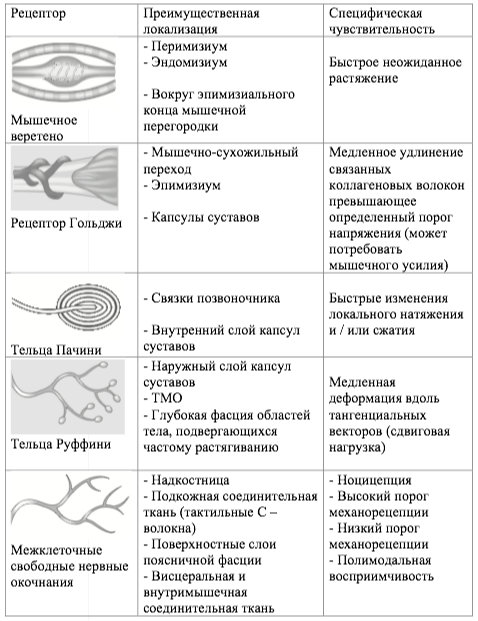
В Таблице 1 приведен обзор различных сенсорных рецепторов в фасции, на которые можно воздействовать с помощью мануальной терапии.
Фасция как мощный генератор боли
То, что фасциальные нервные окончания могут быть причиной боли в мягких тканях, было убедительно показано Tesarz et al. (2011). Это исследование подтвердило наличие свободных нервных окончаний с четкой ноцицептивной функцией в поясничной фасции грызунов и человека. Это также было подтверждено различными провокационными тестами путем введения гипотонического физиологического раствора в поясничную фасцию человека (Schilder et al., 2014). Интересно, что было обнаружено, что связанные с этим интерстициальные нейроны в фасции, по-видимому, особенно чувствительны, с точки зрения последующей длительной гиперчувствительности, к повторному механическому или биохимическому раздражению. Подобные провокационные тесты с гипотоническим солевым раствором показали, что большая часть ощущений, известных как отстроченная мышечная боль (DOMS) после усердных эксцентрических упражнений, по-видимому, происходит благодаря ноцицептивным интерстициальным нейронам в фасциальном слое мышечного эпимизия (Gibson et al., 2009). Новая точка зрения на то, что фасция является потенциальным генератором боли, может иметь важное значение для понимания и лечения боли в пояснице. В то время как некоторые случаи боли в пояснице определенно вызваны деформациями дисков позвоночника, несколько крупных МРТ исследований ясно показали, что для большинства случаев боли в пояснице, возможно, следует искать источники в других частях тела, так как изменения дисков часто являются случайной находкой (Jensen et al., 1994; Sheehan, 2010). Исходя из этого, Panjabi (2006) предложила новую гипотетическую модель объяснения боли в пояснице, которая впоследствии была разработана другими авторами (Langevin & Sherman, 2007; Schleip et al., 2007). Согласно этим авторам, микротравмы в поясничных соединительных тканях могут привести к ноцицептивной передаче сигналов и дальнейшим последующим эффектам, связанным с болью в пояснице. Микротравмы поясничной фасции человека действительно были задокументированы у пациентов с болью в пояснице давно (обзор см. Willard et al, 2012). Однако их присутствие в сопоставимых здоровых людях не было определено количественно, и поэтому причинно-следственная связь оставалась сомнительной. Тем не менее, недавно обширное ультразвуковое исследование Ланеви и Шермана сравнило поясничную фасцию у пациентов с хронической болью в пояснице с группой сопоставимых здоровых людей. Здесь было показано, что этот фасциальный слой более плотно (или более сильно) связан с основной мускулатурой у пациентов с болью по сравнению со здоровой контрольной группой. Этот вывод вместе с ранее опубликованными исследованиями иннервации позволяет предположить, что поясничная фасция должна быть признана в качестве важного фактора для понимания и лечения по крайней мере некоторых случаев боли в пояснице Langevin & Sherman (2007).
Полимодальные рецепторы в фасции
Важно понимать, что не все свободные нервные окончания могут быть классифицированы как ноцицептивные. Некоторые из них являются сенсорными устройствами для терморецепции. Другие сообщают об изменениях мышечной активности в симпатической нервной системе, чтобы обеспечить локальную специфическую настройку кровотока к соответствующим участкам мышц, известную как эргорецепция. Интересно, что в фасциальных тканях большинство интерстициальных нейронов являются так называемыми рецепторами «широкого динамического диапазона» или полимодальными рецепторами, что означает, что они реагируют на более чем один вид стимуляции. (Sandkühler, 2009). Хотя соответствующие им синапсы в заднем роге спинного мозга голодны и жаждут «любого» вида стимуляции, похоже что они легко удовлетворяются, если достаточное количество проприоцептивной информации поступает им через эти полимодальные рецепторы. Однако в случаях недостаточного поступления проприоцептивной стимуляции (например, из-за изменений в матрице соединительной ткани, окружающей соответствующие нервные окончания) эти нейроны имеют тенденцию активно снижать свой порог для ноцицептивной стимуляции. Кроме того, они могут активно выделять цитокины, которые сенсибилизируют полимодальные нейроны по соседству и предрасполагают их к ноцицептивной функции. Кажущаяся минимальной механическая стимуляция, такая как разница в длине ног всего в 1 мм, может затем привести к ноцицептивному ответу в сложной сети этих внутрифазных полимодальных рецепторов. Основываясь на взаимно ингибирующей динамике между проприоцептивной и ноцицептивной интрафасциальной стимуляцией, многие терапевтические подходы исследуют использование движения и / или прикосновения, чтобы снабдить соответствующие полимодальные рецепторы новым проприоцептивным вводом. Это можно сделать разными способами, будь то направленное исследование новых моделей движения (как в методе Фельденкрайза), использование активных микродвижений клиента только с амплитудой в несколько миллиметров (как в Continuum Movement) или использование штангенциркуля для улучшения распознавания двух точек при сближающимся двухточечном прикосновении к коже, параллельно с другими множественными прикосновениями (Buchanan & Ulrich, 2001). По-видимому, индукция нового ноцицептивного ввода - из немного разных мест в пределах одного фасциатома - также может снизить предыдущую гиперчувствительность полимодальных рецепторов. Делать это на хорошо иннервируемой надкостнице (или на переходах сухожилия-надкостницы), по-видимому, особенно эффективно в этом отношении, что может быть одним из объяснительных механизмов, стоящих за более энергичными процедурами, такими как массаж Чуа Ка, и за несколькими аспектами «Модели искажения фасции» Типальдо (Typaldos, 1999).
Интероцептивная иннервация
Несколько свободных нервных окончаний в фасции не связаны ни с соматосенсорной, ни с ноцицептивной передачей сигналов. Вместо этого они информируют островковую кору переднего мозга о физиологических состояниях тканей, таких как тепло, изменения pH или висцеральные ощущения (Craig, 2002). Особое богатство таких интероцептивных окончаний находится в висцеральных соединительных тканях и составляет так называемый кишечный мозг. Интересно отметить, что некоторые дисфункции, знакомые для остеопатов, скорее связаны с нарушением интероцепции, чем с проприоцепцией. Список этих состояний включает синдром раздраженного кишечника, расстройства пищевого поведения, тревогу, депрессию, алекситимию (эмоциональную слепоту) и, возможно, фибромиалгию. Можно предположить, что лечение этих состояний может принести пользу от более интероцептивного, а не проприоцептивного фокуса внимания пациента во время лечения. С другой стороны, лечение состояний с известным проприоцептивным нарушением может лучше всего работать при усиленной проприоцептивной стимуляции и внимании; который затем включал бы боль в пояснице, сколиоз, фантомную боль, хлыстовую травму и комплексный региональный болевой синдром (CRPS) (Schleip et al., 2012). Недавнее открытие «тактильных С-волокон» в подкожных соединительных тканях кожи человека пролило новый свет на мощную и сложную динамику груминг-поведения у приматов. Стимуляция этих нервных окончаний может обеспечить важную интероцептивную передачу сигналов в островковую кору, что приводит к последующему влиянию на психоэндокринную функцию, иммунную систему и вегетативную регуляцию (Schleip et al., 2012).
РЕЗЮМЕ
• Прикосновение к фасции живого человека требует другого подхода нежели к безжизненному куску мяса. Фактически: фасция представляет собой ткань всего тела, которая служит нашим самым большим сенсорным органом. Это обеспечивает нашу самую важную ориентацию в ощущении нашего физического я.
• Участвующие в этом чувствительные нервы включают рецепторы, которые передают проприоцептивную информацию. Различные мануальные методы могут хорошо работать, обеспечивая свежую проприоцептивную стимуляцию этим рецепторам, что приводит к специфическим для рецепторов нервно-мышечным и физиологическим эффектам.
• Фасция также может быть источником боли, в том числе в некоторых случаях боли в пояснице, передаваемой через ноцицептивные нервные окончания.
• Другие нервные окончания в фасции выполняют интероцептивную функцию и имеют тенденцию влиять на островковую кору и ее соматоэмоциональную функцию самочувствия.
• Широко распространенное существование полимодальных рецепторов в фасции создает основу для понимания полезных анальгетических эффектов некоторых фасциальных терапевтических подходов.
ЛИТЕРАТУРА
Buchanan P. A., Ulrich B. D. (2001) The Feldenkrais Method: a dynamic approach to changing motor behavior. Res Q Exerc Sport 72(4):315–323.
Chaudhry H. Schleip R., Ji Z. et al., (2008) Three- dimensional mathematical model for deformation of fascia in manual therapy. J Am Osteop Assoc 108:379–390.
Craig A. D. (2002) How do you feel? Interoception: the sense of the physiological condition of the body. Nat Rev Neurosci 3(8):655–666.
Erdemir A., Piazza S. J. (2004) Changes in foot loading following plantar fasciotomy: a computer modeling study. J Biomech Eng 126(2):237–43.
Findley W. T. & Schleip R. (eds) (2007) Fascia Research: Basic Science and Implications for Conventional and Complementary Health Care. Munich: Elsevier Urban & Fischer.
Gibson W., Arendt-Nielsen L., Taguchi T. et al., (2009) Increased pain from muscle fascia following eccentric exercise: animal and human findings. Exp Brain Res 194(2):299–308.
Jami A. (1992) Golgi tendon organs in mammalian skeletal muscles: functional properties and central actions. Physiol Rev 72(3):623–666.
Jensen M. C., Brant-Zawadzki M. N., Obuchowski N. et al., (1994) Magnetic resonance imaging of the lumbar spine in people without back pain. NEJM 331(2):69–73.
Johansson H., Sjölander P., Sojka P. (1991) A sensory role for the cruciate ligaments. Clin Orthop Relat Res 268:161–178.
Jones T. A. (2004) Rolfing. Phys Med Rehabil Clin N Am 15(4):799–809.
Langevin H. M., Sherman K. J. (2007)
Pathophysiological model for chronic low back pain integrating connective tissue and nervous system mechanisms. Med Hypotheses 68(1):74–80.
Lederman E. (2005) Harmonic Technique. Churchill Livingstone, Edinburgh.
Liptan G. L. (2010) Fascia: a missing link in our understanding of the pathology of fibromyalgia. J Bodyw Mov Ther 14(1):3–12.
Mitchell J. H., Schmidt R. F. (1977) Cardiovascular reflex control by afferent fibers from skeletal muscle receptors. In: Shepherd J. T. Abboud F. M. (eds) Handbook of Physiology, Section 2, Vol. III, Part 2, pp. 623–658.
Panjabi M. M. (2006) A hypothesis of chronic back pain: ligament subfailure injuries lead to muscle control dysfunction. Eur Spine J 15(5):668–767.
Sakada S (1974) Mechanoreceptors in fascia, periosteum and periodontal ligament. Bull Tokyo Med Dent Univ 21 Suppl:11–13.
Sandkühler J. (2009) Models and mechanisms of hyperalgesia and allodynia. Physiol Rev 89(2):707–758.
Schilder A., Hoheisel U., Magerl W., et al., (2014) Sensory findings after stimulation of the thoracolumbar fascia with hypertonic saline suggest its contribution to low back pain. Pain 155(2):222–231.
Schleip R. (2003) Fascial plasticity—a new neurobiological explanation. Part 1. J Bodyw Mov Ther 7(1):11–19.
Schleip R. (2012) Chapter 2.1 Fascia as an organ of communication. In:Schleip R., Findley T. W., Chaitow L., Huijing P. A. (eds.) Fascia: the Tensional Network of the Human Body. Churchill Livingstone Elsevier, pp.77–79.
Schleip R., Naylor I. L., Ursu D., et al., (2006) Passive muscle stiffness may be influenced by active contractility of intramuscular connective tissue. Med Hypotheses 66(1):66–71.
Schleip R., Vleeming A., Lehmann-Horn F., Klingler W. (2007) Letter to the Editor concerning “A hypothesis of chronic back pain: ligament subfailure injuries lead to muscle control dysfunction” (M. Panjabi).
Eur Spine J 16(10):1733–1735.
Stecco C., Gagey O., Belloni A. et al., (2007) Anatomy of the deep fascia of the upper limb. Second part:study of innervation. Morphologie 91(292):38–43.
Stecco C., Porzionato A., Lancerotto L. et al., (2008)
Histological study of the deep fasciae of the limbs. J Bodyw Mov Ther 12(3):225–230.
Still A. T. (1902) The Philosophy and Mechanical Principles of Osteopathy. Hudson-Kimberly Publishing Company, Kansas City, p. 62.
Stillwell D. L. (1957) Regional variations in the innervation of deep fasciae and aponeuroses. Anat Rec 127:635–648.
Tesarz J., Hoheisel U., Wiedenhöfer B., Mense S. (2011) Sensory innervation of the thoracolumbar fascia in rats and humans. Neuroscience 194:302–308.
Trager M., Guadagno-Hammond C., Turnley Walker T. (1987) Trager Mentastics: Movement as a Way to Agelessness. Station Hill Press, Barrytown.
Typaldos S. (1999) Orthopathic Medicine: The Unification of Orthopedics with Osteopathy through the Fascial Distortion Model. Orthopathic Global Health Publications.
van der Wal J. C. (1988) The organization of the substrate of proprioception in the elbow region of the rat [PhD thesis]. Maastricht, Netherlands: Maastricht University, Faculty of Medicine.
van der Wal J. (2009) The architecture of the connective tissue in the musculoskeletal system: an often overlooked functional parameter as to proprioception in the locomotor apparatus. In: Huijing P. A. Hollander P., Findley T. W., Schleip R. (eds), Fascia Research II: Basic Science and Implications for Conventional and Complementary Health Care. Munich: Elsevier GmbH.
Willard F. H. , Vleeming A., Schuenke M. D. et al., (2012) The thoracolumbar fascia:anatomy, function and clinical considerations. J Anat 221(6):507–36.
Yahia L., Rhalmi S., Newman N., Isler M. (1992)
Sensory innervation of human thoracolumbar fascia: an immunohistochemical study. Acta Orthop Scand 63(2):195–197.
Grigoriy Basov ● 16.09.2020 г.
Запись на консультацию и прием по телефону:
+7-909-155-52-38 (Эллада)

Подпишитесь на мой Telegram канал
